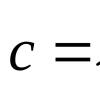Назад
Вперёд
Назад
Вперёд
Внимание! Предварительный просмотр слайдов используется исключительно в ознакомительных целях и может не давать представления о всех возможностях презентации. Если вас заинтересовала данная работа, пожалуйста, загрузите полную версию.
Цель урока: сформировать у школьников понятие о высших споровых растениях, проследить усложнение их организации.
Оборудование . Гербарий мхов, плаунов, хвощей и папоротников, таблицы и рисунки растений, электронный учебник "Биология, 6 класс. Растения. Бактерии. Грибы. Лишайники" издательство "Вентана-Граф", презентация "Высшие споровые растения"
Ход урока
1. Сообщение темы, задач урока и мотивация учебной деятельности
2. Проверка домашнего задания
3. Актуализация опорных знаний и мотивация учебной деятельности
4. Изучение нового материала
1. Предпосылки выхода растений на сушу
Рассказ учителя
Первые растения на Земле появились в воде. Это были водоросли. Водоросли существовали в воде уже миллиарды лет, когда начали. Осваивать сушу первые наземные растения. Почему же растения не могли появиться на суше раньше?
1.Дело в том, что выйти из воды растениям мешали космические лучи, которые убивали все живое. Водоросли могли жить в водоемах, потому что от этих лучей их защищал слой воды. Когда в воде широко распространились фотосинтезирующие водоросли, кислород стал не только накапливаться в воде, но часть его выходила из воды в атмосферу, а из атмосферы в воду переходил углекислый газ, необходимый для фотосинтеза. В результате этих процессов изменился состав атмосферы: она обогатилась кислородом.
2. Особенно важным было то, что из кислорода в атмосфере образовался озоновый слой, который не пропускал на землю смертоносные короткие ультрафиолетовые лучи. В связи с этим стал возможным выход растений на сушу.
2. Приспособления растений к наземно-воздушной среде
Рассказ учителя
Растениям, которые покинули привычную водную среду обитания, нужно было приспособиться к новым условиям жизни на суше, которые резко отличались от привычных. Им нужно было защищаться от высыхания, закрепляться в почве, приспосабливаться к поглощению минеральных веществ и воды из почвы, а кислорода и углекислого газа - из воздуха. Кроме того, растениям необходима была опора, так как воздух, в отличие от воды, не мог их поддерживать.
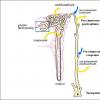
Постепенно растения приспособились к наземному образу жизни. У них появились корни, которые закрепляли растения в почвы и всасывали из нее воду и минеральные вещества. Наземные части растений превратилась в стебель с листьями.
Таким образом, у растений появились органы. Покровные ткани защищали растения от высыхания, а устьица в них осуществляли газообмен для дыхания и фотосинтеза. Возникли проводящие ткани, которые транспортировали воду и минеральные вещества в стебель, и листья, а органические вещества из листьев - по всему растению. И, наконец, образовались механические ткани.
Все эти изменения у вышедших на сушу растений происходил и постепенно.
Первые наземные растения обычно жили во влажных условиях, а их ткани и органы были еще несовершенны. Прошли миллионы лет, пока растения приобрели строение современных представителей растительного мира и заселили все уголки суши. Однако и сейчас еще существуют группы растений с не вполне совершенным строением и значительной зависимостью от воды.
3. Характеристика высших споровых растений
Рассказ учителя
У высших растений в отличие от низших - водорослей, тело делится на органы: корни, стебли, листья. Каждый орган выполняет свои определенные функции. Органы состоят из тканей, которые обеспечивают выполнение этих функций.
В жизненном цикле высших растений наблюдается четкое чередование поколений - спорофита и гаметофита. Гаметофит образует многоклеточные половые органы, в которых развиваются гаметы. При слиянии гамет получается зигота, из которой вырастает следующее поколение - спорофит. На спорофите созревают споры. Споры у растений могут быть все одного или разных размеров - мелкие (микроспоры) и крупные (мегаспоры).
Растения, имеющие одинаковые споры, получили название равноспоровых, а имеющие микроспоры и мегаспоры, - разноспоровых. У равноспоровых из спор вырастают обоеполые гаметофиты. У разноспоровых из микроспоры развивается мужской гаметофит, а из мегаспоры - женский. У высших растений одно поколение по размерам всегда больше другого, и на его долю приходится большая часть жизненного цикла растения. Только у растений одного отдела Моховидные преобладает поколение гаметофитов, у всех остальных высших растений - поколение спорофитов.
Высшие растения делят на две группы - высшие споровые, споровые размножаются спорами, и семенные растения, которые размножаются семенами. Споровые - более древние наземные растения, чем семенные. Они имеют все признаки высших растений, у них не всегда достигают полного развития ткани и органы, пример, проводящая ткань ксилема у всех высших споровых состоит из удлиненных мертвых клеток, а не из сосудов. Еще одной особенностью споровых является то, что размножение, как и у водорослей, связано с водой. Вода необходима для движения мужских гамет - сперматозоидов, имеющих жгутики, неподвижным женским гаметам - яйцеклеткам. При этом гаметы используют воду дождей и россы. Поэтому высшие споровые растения распространены во влажных местах.
4. Классификация высших споровых растений
- Хвощеобразные
- Плаунообразные
- Мохообразные
- Папоротникообразные.
В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
Высшие споровые растения после выхода на сушу в ходе эволюции претерпевали метаморфозы в двух направлениях. Так сформировались две крупные эволюционные группы – гаплоидная и диплоидная. К первой ветви относятся мхи, у которых лучше развит гаметофит, а спорофит занимает подчиненное положение. К диплоидной ветви принадлежат папоротники, хвощи и плауны. У них гаметофит редуцирован, и выглядит как заросток.
Половые органы развиваются на половом поколении. Мужские половые органы - антеридии - образование овальной формы, внутри которых развиваются подвижные сперматозоиды (жгутиковые мужские половые клетки) женские половые органы - архегонии, колбоподибнои формы, у них развивается неподвижная женская половая клетка - яйцеклетка. Для оплодотворения сперматозоид должен попасть во внешнюю среду и оплодотворить яйцеклетку, что находится внутри архегонии. Для перемещения сперматозоида необходима вода. С оплодотворенной яйцеклетки образуется зародыш. Он прорастает и превращается во взрослую особь неполового поколения (спорофит), которая размножается спорами, которые образуются на ней в спорангиях. Особи полового и неполового поколения очень отличаются между собой по строению.
Распространены они в разных климатических зонах. Однако большинство живет на влажных участках суши, хотя некоторые виды этих растений встречаются и в сухих борах, и даже в пустынях.
Отдел Моховидные. В этот отдел входит более 25 тыс. видов. Корней нет. Спорофит самостоятельно не существует, развивается и всегда находится гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий на ножке. Отдел включает класс листостебельные мхи, класс печеночники и антоцеротовые.
Отдел Плауновидные – очень древняя группа, к которой относятся как ископаемые, так и ныне живущие растения. Имеют длинные стелющиеся дихотомически ветвящиеся побеги, густо усаженные жесткими мелкими листьями.
Отдел хвощевидные. Многолетние травы лугов, болот, лесов и полей. От корневищ отходят придаточные корни и надземные побеги, имеющие типичное членистое строение. От узлов отходят мутовки бурых чешуйчатых листьев, сросшихся в трубчатое влагалище, и мутовки боковых побегов.
Отдел папоротниковидные. Преобладание долговечного листостебельного спорофита над эфемерным примитивным гаметофитом, наличие у спорофита крупных, обычно перисто-рассеченных, сложно устроенных листьев – вай, расположение спорангиев группами (сорусами) на нижней стороне листьев.
5. Обобщение, систематизация знаний и умений учащихся
Работа в тетради
Выполнение заданий, предложенных к теме данного урока. Работа с учебником
Школьники читают текст параграфа и отвечают на вопросы учителя.
Вопросы для повторения и обсуждения
- С какими трудностями столкнулись растения, выйдя на сушу
- Почему эти растения относят к высшим растениям?
- Но почему мы добавляем слово споровые растения?
- По каким признакам высшие растения делятся на группы?
- Расскажите об особенностях полового размножения высших споровых растений.
- При каких условиях растения вышли на сушу?
- Как и почему изменился состав атмосферы Земли?
- Перечислите, какие приспособления были нужны растениям, чтобы освоить сушу.
- Почему мхи - тупиковая ветвь эволюции?
- Однажды при разработке торфа был найден хорошо сохранившийся рыцарь в доспехах. Как это можно объяснить?
- Что такое торф? Как его можно использовать?
- В чем проявляется усложнение в строении плаунов?
- В чем проявляется усложнение в строении хвощей?
Ученица зачитывает отрывок из повести Н. Гоголя "Вечера накануне Ивана Купала".
Петро, герой повести Н. Гоголя "Вечер накануне Ивана Купала", так увидел цветение папоротника: "Глядь, краснеет маленькая цветочная почка и, как будто живая, движется. В самом деле, чудо! Движется и становится все больше, больше и краснеет, как горячий уголь. Вспыхнула звездочка, что-то тихо затрещало, и цветок развернулся перед его очами, словно пламя, осветив и другие около себя. "Теперь пора!" - подумал Петро и протянул руку. Смотрит, тянутся из-за него сотни мохнатых рук также к цветку, а позади его что-то перебегает с места на место. Зажмурив глаза, дернул он за стебелек, и цветок остался в его руках".
Учитель: Так что же из себя представляет папоротник?
6. Подведение итогов урока
7. Домашнее задание.
Изучить материал учебника к теме урока, выполнить задания в рабочей тетради.
Споровые растения - растения, размножающиеся и распространяющиеся спорами, которые образуются бесполым или половым путем. К Споровым растениям относят мхи, плауны, хвощи, папоротники.
Споровые растения еще называют архегониальными . Тело высших растений дифференцировано на ткани и органы, которые появились у них как одно из приспособлений к жизни на суше. Наиболее важные органы - корень и побег , расчлененный на стебель и листья. Кроме того, у наземных растений формируются специальные ткани: покровная , проводящая и основная .
Покровная ткань выполняет защитную функцию, защищая растения от неблагоприятных условий. Через проводящую ткань проходит обмен веществ между подземными и надземными частями растения. Основная ткань выполняет различные функции: фотосинтезирующую, опорную, запасающую и т. д.
У всех споровых растений в их жизненном цикле развития четко выражено чередование поколений: полового и бесполого.
Половое поколение - заросток, или гаметофит - образуется из спор, имеет гаплоидный набор хромосом. Оно выполняет функцию образования гамет (половых клеток) в специальных органах полового размножения; архегониях (от греч. «архе» - начало и «гоне» - рождение) - женских половых органах и антеридиях (от греч. «антерос» - цветущий) - мужских половых органах.
Ткань спорангиев также имеет двойной набор хромосом, она делится путем мейоза (способ деления), в результате чего развиваются споры - гаплоидные клетки с одинарным набором хромосом. Название поколения «спорофит» означает растение, образующее споры.
У мхов преобладает гаметофит (половое поколение) у хвощей, плаунов, папоротников - спорофит (бесполое поколение).
Моховидные, или мхи, представляют собой обособленную группу высших растений, развитие которой привело к эволюционному тупику. В отличие от всех других отделов высших растений, в жизненном цикле мхов гаплоидный гаметофит преобладает над спорофитом и осуществляет функции фотосинтеза, обеспечения водой и минеральным питанием.
Риччия - часто встречающееся в аквариумах растение. Представляет собой ажурный сочно-зеленый мох, плавающий по поверхности воды и образующий очень красивые островки. У этого растения нет ни стеблей, ни листьев, ни корней. Она состоит из мелких ветвящихся плоских пластинок, так называемого слоевища.
Мох ключевой. Обычно мох ключевой произрастает большими группами, прикрепляясь к камням на дне водоема. Сильно разветвленные стебли покрыты многочисленными листочками длиной около 1 см и шириной 0,5 см. Цвет растения зависит от места обитания и колеблется от светло-зеленого до темно-зеленого.
Мох яванский. Длинны, сильно ветвящиеся стебли достигают 50 см. Представляет собой переплетение тонких нитей темно-зеленого цвета, маленькие (около 0,2 см) листочки окрашены в различные оттенки зеленого цвета.
Якорный мох. Медленно растёт при любом освещении. Мох тонет в воде, при добавке углекислоты формируется пышными кустами.
Мох феникс. Один из водных видов мхов. Произрастает в Мексике. Диапазон освещения: от слабого до очень сильного. Ризоиды этого мха хорошо зацепляются за дерево или камень. Имеет малые размеры и медленный рост.
Печеночный мох- в летнее время, быстро разрастаясь, печеночный мох заполняет всю поверхность воды, препятствуя поступлению в аквариум кислорода из атмосферы, поэтому печеночный мох необходимо периодически удалять. При этом следует оставлять более сильные ветки, ещё их называют рогульки, которые образуются обычно в наиболее освещенном месте аквариума.
Павлиний мох. Растет медленно. Рост можно ускорить, если повысить уровень освещенности. Также немаловажным условием для его более интенсивного роста является температура воды, не превышающая 25°С. Если увеличить температуру до 30°С, то мох листья мха начнут деформироваться.
(12)ПЛАМЯННЫЙ МОХ
(13)ПЛАЧУЩИЙ МОХ
(14)ИЗОГНУТЫЙ МОХ
(15)БЛЕФАРОСТОМА
(16)КИТАЙСКИЙ МОХ

Плауновидные – древние растения, произошедшие от риниофитов, по-видимому, в середине девонского периода палеозойской эры и достигшие расцвета в каменноугольном периоде. В то время существовали гигантские формы плаунов. Одни из популярных сегодня видов аквариумных растений – папоротники. Это споровые растения, которые могут самостоятельно развивать и размножаться в случае подходящих условий. Также некоторые виды могут расти в разных температурных условиях, что делает этот вид растений практичными. Внешне все виды папоротников похожи между собой, но различить их реально. Они относятся к самой большой группе растений, размножающихся спорами.
(18) Плаун баранец
(19) Плаун куропаточий.
(20) Плаун сплюснутый
(21) Плаун можжевельниковый
(22) Плаун годичный
(23) ХВОЩИ
Хвощевидные – немногочисленная группа растений, насчитывающая около 20 видов. Гораздо шире они были представлены в позднем девонском и каменноугольном периодах
(24)ХВОЩ ЗИМУЮЩИЙ
(25) Хвощ приречный
(26)ПАПОРОТНИКОВЫЕ
Папоротниковидные, или папоротники, как и другие высшие споровые растения, произошли от риниофитов в девоне и достигли расцвета в каменноугольном периоде палеозойской эры.
Азолла каролинская или водяной папоротник
Азолла каролинская – это водное растение, которое не растет в глубине воды, а плавает на ее поверхности. На ее листьях растут некие водоросли, которые способствуют поглощение азота и кислорода. Так растение «питается». Несколько растений азоллы могут создать на поверхности воды зеленые участки, будто ковры.Растение очень нежное, требует бережного обращения. Встречается у аквариумистов сравнительно редко. Имеет выраженный сезонный характер роста с периодом покоя в зимнее время.
(28) Вольфия бескорневая
К температурным условиям растение не требовательно.
Его можно выращивать в умеренно теплом и тропическом аквариумах.
(29) Лимнобиум побегоносный Лимнобиум представляет собой плавающее по поверхности растение с круглыми глянцевыми листьями диаметром 2-3 см, сидящими на коротких черенках. Используется не только как декоративное растение, но и в качестве естественного затенителя в аквариуме. При благоприятных условиях лимнобиум, быстро разрастаясь, затягивает всю поверхность аквариум
(30) Пистия или Водяной салат
Пистия - одно из самых красивых плавающих на поверхности воды растений. Представляет собой розетку крупных бархатистых листьев голубовато-зеленого цвета. В диаметре розетка может достигать 25 см. Крупные экземпляры растения достигают высоты 15 см. Пистия имеет хорошо развитую корневую систему, состоящую из множества длинных корешков. Переплетение корней могут опускаться на значительную глубину, до 25-30 см.
РЯСКА МАЛАЯ
Состоит из отдельных листьев округлой формы светло-зеленого цвета диаметром до 5 мм. Тонкие нитевидные корешки могут иметь длину до 10 см.
САЛЬВИНИЯ ПЛАВАЮЩАЯ
Растение имеет короткие стебли, на которых попарно расположены ярко-зеленые листья длиной до 1,5 см, округлой формы, покрытые снизу тонкими бурыми волосками
(33)САЛЬВИНИЯ УШАСТАЯ
Стебель ветвящийся, короткий. Листорасположение мутовчатое по 3 листочка в мутовке. Два плавающих листа от округлой до продолговатой формы, находятся напротив друг друга и имеют по две выпуклости, покрыты короткими волосками, воды касаются только краями и средней жилкой. Третий листок опущен вниз, нитевидно расчленен и похож на корень. Окрас листьев от светло-зеленого до голубовато-зеленого цвета.
(34) Людвигия ползучая
(35)ПАПОРОТНИК РОГАТЫЙ
(36) Папоротник индийский водяной
(37) Папоротник таиландский, крыловидный
(38) Ротала круглолистная или Ротала индийская
(39) Лимнофила водная, амбулия водная
(40) Апоногетон Капурони
(41)Аир травянистый (акорус)
(42)Гидрокотила белоголовая или Щитолистник белоголовый
Подцарство высших растений включает в себя зеленые растения, обитающие на суше. В зависимости от степени дифференци- ровки тканей и особенностей размножения их делят на две неравные но величине и значению группы - высшие споровые растения и семенные растения. Споровые растения размножаются и расселяются с помощью спор. Семенные растения морфологически более сложно устроены, и единицей размножения и расселения у них служит семя. В подцарстве высших известны миогочисленные ископаемые виды растений. Современные представители объединены в пять отделов споровых и два отдела семенных растений.
Высшие споровые растения
Высшие споровые растения населяют сушу более 400 млн лет. Первые растения были небольшого размера, имели просто устроенные вегетативные органы. В процессе эволюции совершенствовалось внутреннее и внешнее строение. У высших споровых растений споры образуются в многоклеточных спорангиях и приспособлены к распространению ветром. Из спор развивается гаметофит, который называется заросток, на нем формируются половые органы. Для полового размножения необходима капельно-жидкая вода. Бесполое и половое поколения у споровых представлены самосто- ятельным и организмами.
К высшим споровым растениям относятся отделы: моховидные, плауновидные, псилотовидные, хвощевидные, папоротниковидные.
Отдел моховидные. К этой группе относятся древнейшие и наиболее просто устроенные высшие растения. Характерной чертой мхов, которая отличает их от всех высших растений, является доминирование в жизненном цикле гаметофита - полового поколения. К другим признакам моховидных относятся следующие:
- у наиболее примитивных представителей тело представлено талломом. Более организованные представляют собой листостебельные растения;
- у моховидных нет корней, к субстрату растения прикрепляются ризоидами;
- у мхов нет проводящей системы, в отличие от всех высших растений мхи - бессосудистые растения;
- поглощение и испарение воды осуществляется через всю поверхность тела;
- у сфагновых мхов развита водозапасающая ткань;
- не развиты механические ткани, поэтому мхи небольшого размера, растут куртинами;
- большинство мхов размножается вегетативно (с помощью частей веточек, листьев, выводковых почек).
Зеленое растение мхов представлено гаметофитом, на котором образуются гаметангии. Они формируются на верхушке побега. Оплодотворение возможно только при наличии воды. В каплю воды выделяется множество сперматозоидов и вместе с ней может переноситься с растения на растение. После оплодотворения развивается спорофит. Он представляет собой коробочку с ножкой, с помощью которой присасывается к зеленому растению-гамето- фиту и живет за счет него. Спорофит и гаметофит на этом этане представляют как бы одно растение. В коробочке созревает множество спор, они рассыпаются и переносятся ветром. Среди мхов имеются как однодомные, так и двудомные организмы.
Мхи - многолетние небольшие вечнозеленые растения. В течение жизни они могут полностью высыхать, но после смачивания возвращаться к вегетации. Мхи очень медленно растут, всего несколько миллиметров в год. Сфагновые мхи известны как природные антисептики, они мало поддаются гниению и формируют торфяные залежи на верховых болотах. Большое значение имеют мхи для водообмена - они задерживают атмосферную воду и переводят ее в состояние грунтовой. Из-за особенностей строения мхи заселяют наиболее влажные местообитания. Они встречаются в тундре, в таежных лесах, на верховых и переходных болотах. Встречаются луговые и эпифитные мхи.
Моховидные достаточно разнообразны (рис. 9.21). Среди них встречаются плоские слоевищные растения, такие как маршанция (класс печеночники) - напочвенное растение в виде дихотомически ветвящейся веточки размером до 10 см. Маршанцию можно встретить на влажных местах, в лесу на старых кострищах, на древесине.
Рис. 9.21.
- 1 - политрихум обыкновенный, или кукушкин лен (Polytrichum commune, а - лист, б - коробочек); 2 - маршанция многообразная (Marchantia );
- 3 - мний (Mnium utidulatum ); 4 - сфагнум (Sphagnum )
Сфагновые мхи распространены по все миру, заселяют верховые болота, образуют на них большие подушковидные куртины. Сфагнумы - растения бледно-зеленого цвета, высыхая, становятся белесыми или светло-бурыми, за что их называют белыми мхами.
Сфагновые мхи могут быстро и в большом количестве накапливать воду в мертвых водоносных клетках.
Самая многочисленная группа мхов - бриевые или настоящие мхи. Широко распространен мох политрихум обыкновенный, или кукушкин лен, обитатель хвойных лесов, сырых лугов, болот. Его стебель, до 40 см длиной, густо прокрыт жесткими узкими листьями. В таежных лесах политрихум образует сплошной покров, такие леса называют долгомошными.
Отдел плауновидные. К этой группе относятся растения с развитыми проводящими тканями. Вместе с хвощевидными и папоротниковидными они составляют группу сосудистых споровых растений.
Плауны одна из наиболее древних групп растений. В каменноугольный период плауновидные представляли собой огромные древовидные формы. Ископаемые лепидодендроны были высотой до 30 м. Они и другие вымершие споровые растения сформировали залежи каменного угля.
Современные нлауновидные - это многолетние вечнозеленые травы, высотой до 15-20 см. У плаунов хорошо развиты вегетативные органы. Плауны имеют длинный ползучий дихотомически ветвящийся стебель, покрытый жесткими мелкими чешуевидными листочками. От побега отрастают придаточные корни. Вертикальные веточки заканчиваются спороносными колосками (рис. 9.22). Созревшие в них споры разносятся ветром и в благоприятных условиях прорастают в очень маленькие (2-3 мм) заростки. Бесцветный гамето- фит развивается под землей в течение 15-20 лет. Заростки способны существовать только благодаря симбиотическим грибам. На заростках образуются гаметы, и в присутствии воды происходит оплодотворение, после чего развивается новое растение-спорофит. Плауны могут размножаться и вегетативно, частями стебля.
Споры плауна содержат много масла. Раньше они использовались в пиротехнике, для изготовления мелких и фасонных форм для литья (художественное литье в Касли), как детская присыпка, для обсыпки пилюль.
В современной флоре нлауновидные занимают скромное положение. Главные представили отдела - виды рода плаун, связаны с хвойными лесами Азии и Северной Америки (плаун годичный, плаун булавовидный, баранец). В более теплых местообитаниях распространены мелкие растения рода селягинелла. Она интересна тем, что у нее формируется два вида спор - микро- и мегаспоры, из которых развиваются разнополые заростки. Это новое для наземных растений явление - разноспоровость, разовьется у более эволюционно продвинутых групп.

Рис. 9.22. Плаун булавовидный (Lycopodium clavatum):
1 - общий вид спорофита; 2 - спорофилл со спорангием; 3 - спора с двух сторон; 4 - заросток; 5 - заросток с молодым спорофитом
Отдел хвощевидные. Это самый малочисленный среди споровых растений отдел. В каменноугольном периоде хвощи были представлены большим разнообразием форм. Вымершие древовидные каламиты образовывали болотные леса. Современные хвощевидные - это многолетние травянистые растения. Из-за особенностей строения вегетативного тела хвощи называют также членистые - побег имеет выраженное метамерное строение. Метамером является полое междоузлие и мутовка боковых веточек. Листья на побеге редуцированы, превратились в мелкие щетинки. Они расположены супротивно на боковых стеблях и образуют мутовку в узлах главного побега. Функция фотосинтеза перешла к зеленым стеблям. Часто покровы хвощей пропитаны кремнеземом, поэтому на ощупь эти растения очень жесткие. От подземного горизонтального корневища отрастают придаточные корни, часто на корневище образуются клубеньки, заполненные крахмалом. Весной от корневища отрастают вертикальные спороносные побеги, они светло-бурого цвета и выполняют только функцию размножения. Летом на этих же корневищах развиваются зеленые вегетативные побеги. У некоторых хвощей спорангии образуются на зеленых побегах, т.е. вегетативная и спороносная функции не разделены. Спорангии расположены в верхушечных спороносных колосках, спорофиллы имеют оригинальное строение в виде шестигранного щитка. Из спор вырастают заростки, которые имеют форму маленьких зеленых пластиночек. Заростки развиваются быстро, и через несколько недель на них развиваются антеридии и архегонии, после оплодотворения начинает развиваться молодой спорофит. Для оплодотворения, как и веем споровым, необходима вода. Растения успешно размножаются вегетативно, частями подземного корневища.
Хвощевидные - это, в основном, растения Северного полушария, обитают на влажной кислой почве в сырых лесах, на болотах, на влажных лугах и полях. В культурных ценозах хвощи являются злостными сорняками. Типичными представителями этого отдела являются хвощ лесной, хвощ топяной, хвощ полевой (рис. 9.23), хвощ приречный.

Рис. 9.23. Хвощ полевой (Equisetum arvense):
- 1 - общий вид спорофита {а - вегетативные побеги с мутовчато расположенными боковыми веточками, б - спороносные весенние побеги);
- 2 - спорофилл - щиток с верхней и нижней стороны; 3 - шаровидная спора с закрученными элатерами; 4 - споры с раскрученными элатерами
Отдел папоротниковидные. Древние ископаемые папоротники, совместно с другими вымершими плаунами и хвощевидными, составляли мощные каменноугольные леса. В современном растительном покрове представители этого отдела самые распространенные из споровых растений. Более всего их во влажных тропиках. Там встречаются древовидные папоротники, высотой до 20 м, травянистые формы, эпифиты и лианы. Папоротники умеренного пояса - это травянистые растения с многолетним подземным корневищем, на котором каждый год образуются пучки новых крупных листьев. Листья имеют характерную особенность - они растут своей верхушкой, как стебель. Неразвившиеся листья сложены улиткообразно, вырастая, они разворачиваются в плоский лист, расчлененный на черешок и листовую пластинку. Такой лист имеет свое название - вайя. У большинства папоротников листья перистые. Некоторые папоротники имеют отдельно вегетативные и репродуктивные листья - спорофиллы.
Зеленое растение папоротника - спорофит. Спорангии у папоротников расположены на нижней стороне листьев, группами, которые называются сорусы. У многих видов сорусы защищены особым покрывальцем, что спасает споры от высыхания. Для папоротников характерны различные приспособления для разбрасывания спор. Споры имеют многослойную оболочку и хорошо переносят неблагоприятные условия, долго сохраняют способность к прорастанию. Из споры развивается заросток - это самостоятельно живущая обоеполая зеленая пластинка размером до 5 мм. На пей развиваются гаметы, с участием воды происходит оплодотворение. Из зиготы развивается новый папоротник.
Папоротниковидные хорошо приспособлены к различным условиям среды, растут по всему земному шару, встречаются в самых разнообразных местообитаниях, чаще во влажных лесах, на болотах, лугах. Широко распространены папоротники в лесах умеренной зоны - это орляк, щитовник (рис. 9.24), кочедыжник, стра- усник и многие другие. Встречаются засухоустойчивые формы, наскальные (из родов вудсия, асплениум, полиподиум) и растущие в воде, например плавающий папоротник сальвиния.

Рис. 9.24.
1 - внешний вид папоротника (бесполое поколение); 2 - долька листа с нижней стороны (видны сорусы, одетые покрывалом); 3 - разрез соруса, а - спорангии, б - покрывало; 4 - отдельный спорангий, из которого
высыпаются споры
Высшие растения.
У большинства высших растений тело дифференцировано на органы - корень, стебель и листья, состоящие из хорошо обособленных тканей. В жизненном цикле высших растений четко выражено чередование спорофита (2n) и гаметофита (n). Органы полового размножения многоклеточные. Женский - архегоний - состоит из расширенной нижней части - брюшка, где формируется яйцеклетка, и верхней суженной - шейки, раскрывающейся при созревании яйцеклетки. Мужской орган полового размножения - антеридий - имеет вид мешка, внутри которого образуется.множество сперматозоидов. У голосеменных антеридии подверглись редукции, а у покрытосеменных редуцированы и антеридии, и архегоний. Из зиготы у высших растений образуется зародыш - зачаток спорофита.
Отдел Моховидные - Bryophyta.
Общее число видов около 35 тыс.
Строение . В жизненном цикле моховидных, как и других высших растений, имеется чередование двух фаз: спорофита и гаметофита. Однако доминирует (преобладает) гаметофит, в то время как у всех других высших растений доминирует спорофит. Именно поэтому моховидные рассматривают как самостоятельную боковую ветвь в эволюции высших растений.
Моховидные по своей организации и экологии еще близки к водорослям. Как и у водорослей, у них нет сосудов и корней. Некоторые примитивные представители имеют вегетативное тело в виде стелющегося таллома с верхушечным (дихотомическим) ветвлением, похожего на таллом водорослей. Оплодотворение связано с водой. Среди моховидных, как и среди водорослей, нет одревесневающих форм.
Распространение . Моховидные распространены на всех континентах мира, но неравномерно. В тропических странах - преимущественно в горах. Незначительное число видов растет в засушливых условиях, например в степях. Некоторые виды ведут эпифитный образ жизни на корке деревьев или водный. Основное же разнообразие видов сосредоточено во влажных местах Северного полушария, в областях с умеренным и холодным климатом. В сложении растительного покрова, особенно тундр, болот и лесов, им принадлежит важная роль.
Классификация . Моховидные подразделяют на три класса: Антоцеротовые, Печеночники, Листостебельные мхи. Наибольшее значение имеют два последних класса.
Класс Печеночники - Hepaticopsida.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности.
Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
Класс Листостебельные мхи - Bryopsida.
Общее число видов около 25 тыс. Многие виды распространены в приполярных странах Северного полушария. На огромных территориях в тундре, на болотах, в лесах они доминируют в растительном покрове, значительно влияя на обеспеченность суши влагой.
Гаметофит представляет собой прямостоячую стеблевидную ось - каулидий, покрытую листовидными выростами - филлидиями. Условно их можно называть стебель и листья. На нижней части стебля образуются многоклеточные ризоиды (не у всех). Ветвление боковое. Нарастание осей происходит в результате деления пирамидальной верхушечной клетки. Оно может быть моноподиальным или симподиальным. В соответствии с этим органы полового размножения и спорогон размещаются на вершине гаметофита или на боковых разветвлениях.
Класс подразделяют на три подкласса: Андреевые мхи, Сфагновые мхи, Бриевые (Зеленые) мхи. Наибольшее значение имеют два последних подкласса.
Подкласс Сфагновые мхи - Sphaqnidae.
Сфагновые мхи имеют довольно однообразное строение и поэтому трудны для определения. Гаметофит их представляет собой сильноветвящееся, особенно в верхней части, растение. Разветвления густо покрыты листьями. Сфагновые мхи живут в очень влажной среде. В связи с этим у них нет ризоидов и влага поступает непосредственно в стебель, который при основании с течением времени отмирает. Строение стебля несложное. В центре его находится сердцевина из тонкостенных паренхимных клеток, выполняющих проводящую и запасающую функции. Она окружена корой, состоящей из двух слоев: склеродермы, выполняющей механическую функцию, и гиалодермы, выполняющей водозапасающую функцию. Клетки гиалодермы крупные, мертвые, стенки их имеют круглые отверстия, через которые полости смежных клеток сообщаются друг с другом, а также с внешней средой. Иногда эти клетки несут спиральные утолщения. Лист состоит из одного ряда клеток, резко различающихся как по структуре, так и по выполняемой функции. Одни из них живые, хлорофиллоносные, другие - мертвые, относительно более крупные, со спирально утолщенными стенками, пронизанные отверстиями, по строению сходные с водозапасающими клетками гиалодермы, их называют гиалиновыми. Гиалиновые клетки способны накапливать и долго удерживать огромное количество воды, в 30 - 40 раз превышающее массу самого растения.
Гаметофиты - однодомные и двудомные. Антеридии образуются в пазухах листьев на разветвлениях стебля. Около них листья окрашены в красноватый цвет. Архегонии на укороченных разветвлениях. В результате слияния сперматозоида с яйцеклеткой возникает зигота, которая представляет собой начало диплоидной фазы - спорогона. Спорогон состоит из ножки и коробочки. Ножка сильно укорочена, луковицеобразная, но ко времени созревания спор верхушка стебля гаметофита сильно вырастает и выносит коробочку кверху (ложная ножка). В центре коробочки помещается округлая колонка, над которой размещен в виде свода спорангий со спорогенной тканью. Стенка коробочки прочная, многослойная. Наружный хлорофиллоносный слой содержит большое число недоразвитых устьиц. Коробочка имеет крышечку, которая во время созревания спор отскакивает, и споры рассеиваются. Элатер нет. Из спор образуется сначала зеленая пластинчатая протонема, а затем из почек, расположенных на ней, - взрослый гаметофит, который и доминирует в жизненном цикле.
Структура сфагновых отличается примитивностью: пластинчатая протонема, отсутствие проводящего пучка и ризоидов, слабая дифференциация коробочки.
Значение сфагновых в природе очень велико. Накапливая огромное количество воды и разрастаясь плотными дернинами, они вызывают заболачивание огромных пространств, достигающих зоны тундры. Для осушения их осуществляют агромелиоративные работы. С другой стороны, старые болота имеют важное хозяйственное значение для разработки залежей торфа. Нарастание пласта торфа в наиболее благоприятных условиях происходит медленно - слой толщиной 1 см образуется примерно за 10 лет.
Подкласс Бриевые (зеленые) мхи - Bryidae.
Число видов 24,6 тыс. Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты.
Отделы Риниовидные - Rhyniophyta и Псилотовидные - Ps1lotophyta.
Отдел Риниовидные включает 2 - 3 рода только ископаемых растений. В жизненном цикле доминирует спорофит. Вегетативное тело его состоит из системы разветвленных теломов. Общая структура в надземной части телома очень своеобразна. Это еще не побег, поскольку листьев на осях телома нет. Главная ось хорошо выражена. Ветвление верхушечное (дихотомическое). В центре оси обособлена ксилема, окруженная флоэмой. Ксилема может быть расположена компактно в виде цилиндра или в виде лучей. Она состоит из трахеид. Периферическая (коровая) часть телома выполняет функцию фотосинтеза. В эпидерме находятся устьичные аппараты. На подземной части устьиц нет. Настоящих корней нет, их заменяют ризоиды. Спорангии расположены на верхушках телома, стенка спорангия многослойна. Гаметофиты риниовидных не найдены. Представитель - род риния (Rhynia), включающий два вида. Это травянистые растения высотой около 20 см, диаметром 3 мм. Подземная часть состоит из горизонтального телома, от которого перпендикулярно отходят воздушные оси.
К отделу Псилотовидные в современной.флоре относят два рода: псилот (Psilotum) и тмезиптер (Tmesipteris). Общее число видов 4 - 6. Оба рода имеют широкое распространение в тропической и субтропической зонах обоих полушарий.
Спорофит псилотовидных - эпифитное, реже наземное травянистое растение. Телом длиной 5 - 40 (до 100) см. Ветвление чаще верхушечное. Кора хорошо развита, выполняет функцию фотосинтеза. Устьичные аппараты примитивные. Листья мелкие, длиной 1 - 5 мм, шиловидные, плоские, без устьичных. аппаратов и жилок. Их можно рассматривать как выросты телома. Подземная часть представлена корневищем с ризоидами. Корней нет. Спорангии срастаются по 2 - 3 (синангии), вскрываются продольной щелью. Споры одинаковой величины. Структура спорофита псилотовидных свидетельствует о близости к риниовидным.
Гаметофит обоеполый, без хлорофилла, радиально симметричный, ветвление верхушечное. Длина его примерно 20 мм, диаметр 2 мм, Питается сапрофитно при помощи грибов, с которыми вступает в симбиоз. Поверхность покрыта ризоидами. Живет чаще под землей. Оплодотворение связано с водой.
По сравнению с риниовидными псилотовидные стоят на более высокой ступени эволюции.
Отдел Плауновидные - Lycopod10phyta.
Плауновидные - одни из наиболее древних высших растений. Они представляют мелколистную линию эволюции. Современные виды - многолетние вечнозеленые травянистые растения, среди вымерших были и древесные формы.
Строение . Спорофит имеет надземный побег с мелкими, иногда чешуевидными листьями (микрофиллами). Они слабо дифференцированы, имеют 1 - 2 неветвящиеся жилки. Узлы и междоузлия выражены слабо. Подземная часть спорофита представлена корневищем с придаточными корнями. Ветвление надземных и подземных осей верхушечное. Спорангии располагаются на верхней стороне листьев (спорофиллов), собранных на концах осей в колоски, реже образующих на стебле спороносные зоны. Споры одинаковой или разной величины.
Гаметофиты подземные, длиной 2 - 20 мм, питаются сапрофитно. Оплодотворение связано с водой.
Классификация . Отдел делят на два класса: Плауновые и Полушниковые.
Класс Плауновые - Lycopodiopsida.
Спорофиты - травянистые многолетние растения. Стебель и корень не имеют камбия. Листья без язычков. Споры одинаковой величины. Гаметофиты обоеполые, созревают в течение 1 - 15 лет. Многие виды вымерли. В современной флоре класс представлен двумя родами. Наиболее многочисленный и широко распространенный из них - род плаун (Lycopodium). Хозяйственное значение плаунов невелико. Животные их не поедают. Плауны служат сырьем для производства лекарств. Издавна используют споры плаунов, содержащие невысыхающее масло. Их применяют в качестве детской присыпки, а иногда при фасонном литье для обсыпания стенок моделей, чтобы отливаемая деталь имела гладкие стенки и легко отделялась от формы.
Плаун булавовидный (Lycopodium clavatum). Растение, широко распространенное в хвойных лесах.
Спорофит представлен длинным ползучим побегом с вертикальными ответвлениями и придаточными корнями. Побег и корни имеют верхушечное ветвление. Проводящий пучок расположен в центре стебля. Центральный цилиндр занимает небольшую часть. Широкая зона коры пронизана листовыми следами. Камбия нет. На поверхности стебля и листьев имеется эпидерма с устьичными аппаратами. Листовая пластинка линейная, цельнокрайняя, заканчивается длинным тонким волоском.
Спороносные колоски, венчающие вертикальные побеги, расположены на довольно длинных ножках по два (реже по 3 - 5). Колосок цилиндрической формы, состоит из оси, на которой плотно сидят спорофиллы - чешуевидные треугольные листья с заостренными и загнутыми кверху верхушками. На верхней стороне спорофилла расположен на короткой ножке почковидный спорангий со спорами. Споры одинаковые, мелкие, тетраэдрической формы. Спородерма имеет два слоя: наружный - экзину, и внутренний - интину. Спорангий растрескивается поперечной щелью.
Споры падают на землю, и на глубине нескольких сантиметров из них медленно, в течение 12 - 15 лет, развивается гаметофит. По форме он напоминает луковичку, позднее разрастается и становится блюдцевидным диаметром до 2 см. Гаметофит бесцветный. Клетки, расположенные под эпидермой, находятся в симбиозе с мицелием гриба. У некоторых видов гаметофит образуется на поверхности почвы, и тогда в его клетках появляются хлоропласты. Антеридии и архегонии размещены на верхней стороне и погружены в паренхимную ткань. Сперматозоиды многочисленные, двухжгутиковые. Оплодотворение связано с водой. Зигота не имеет периода покоя, из нее сразу же образуется зародыш спорофита. Он вначале внедрен в ткань гаметофита и в какой-то мере питается за его счет, но вскоре корни его проникают в почву, и начинается долгая самостоятельная жизнь спорофита.
Класс Полушниковые - Isoetopsida.
Спорофиты представлены деревьями, имеющими вторичное утолщение стебля, и многолетними травами, отчасти сохранившими способность к вторичному утолщению. Древесные виды полностью вымерли. Листья на поверхности, обращенной к стеблю, имеют небольшой вырост - язычок. Споры разной величины. Гаметофиты раздельнополые, созревают в течение нескольких недель. Наиболее многочисленным и широко распространенным является род селагинелла (Selaginella). Используют лишь немногие ее виды как лекарственные и декоративные растения.
Селагинелла селаговидная (Selaginella selaginoides). В нашей стране изредка растет на влажных субальпийских лугах, расположенных в горах европейской части. Спорофит по внешнему виду похож на спорофит плауна. В центре стебля находится проводящий пучок, подвешенный на нитях из паренхимных клеток. Эпидерма стебля без устьичных аппаратов. Листья одинаковые по форме и величине, язычок со временем отваливается. В клетках мезофилла имеется по 1 -2 пластинчатых хлоропласта. Клетки эпидермы также содержат хлоропласты. Устьичные аппараты размещаются чаще на нижней стороне листа. Спороносные колоски расположены на верхушке побегов. Спорофиллы заметно отличаются от вегетативных листьев, они несут язычки, которые не отваливаются. В пазухах спорофиллов каждого колоска сидят на коротких ножках мега- и микроспорангии. В мегаспорангии образуется четыре мегаспоры, в микроспорангии - множество микроспор. Освободившись из спорангиев, мега- и микроспоры в благоприятных условиях прорастают. Внутри микроспоры образуется мужской гаметофит. Он состоит из одной вегетативной клетки и одного крайне редуцированного антеридия, внутри которого образуются двухжгутиковые сперматозоиды. Женский гаметофит также почти целиком формируется внутри мегаспоры. Разрастаясь, гаметофит разрывает покровы мегаспоры, Через трехлучевую щель выдвигается часть тела гаметофита, на которой образуются архегонии и ризоиды. Оплодотворение связано с водой. Из зиготы вырастает зародыш.
Таким образом, разноспоровость обусловила разнополость гаметофитов и их редукцию. Это сыграло важнейшую роль в эволюции растений.
Отдел Хвощевидные - Equisetophyta.
Древовидные представители полностью вымерли, в современной флоре сохранились только травянистые.
Строение . Характерная особенность спорофита - боковое ветвление с мутовчатым расположением боковых побегов. Листья также собраны в мутовки. Узлы и междоузлия четко выражены. При основании междоузлий часто имеется интеркалярная меристема. Листья редуцированные, от средних до мелких, с одной средней жилкой. Спорангии расположены на спорангиофорах - гомологах спорофиллов, но отличающихся от них структурой. Выражен гетероталлизм (физиологическая разноспоровость) - из одинаковых по размеру спор образуются разнополые гаметофиты. Спородерма, кроме интины и экзины, имеет еще наружный слой - перину. Она состоит из двух спирально закрученных лент - элатер, прикрепленных к экзине. Элатеры выполняют двойную функцию: распространения спор и соединения спор в группы, что обеспечивает совместное произрастание мужского и женского гаметофитов.
Гаметофиты мелкие, величиной в несколько миллиметров, зеленые, одно- или (реже) обоеполые. Оплодотворение связано с водой. Зародыш не имеет периода покоя.
Классификация . Отдел подразделяют на четыре класса: Гиениевые, Клинолистные, Каламитовые, Хвощовые. Первые три класса вымерли. Хвощовые представлены в современной флоре обоих полушарий.
Класс Хвощовые- Equisetopsida
Класс включает один род - хвощ (Equisetum). Общее число видов 30 - 35, на территории бывшего СССР - 13. Распространены по всему земному шару, кроме Австралии. Растут преимущественно в условиях влажных местообитаний. По внешнему облику хвощи близки к своим древесным предкам - каламитовым, но значительно меньше их. Надземные побеги у хвощей, как правило, однолетние, лишь у немногих видов - многолетние, вечнозеленые. Отдельные виды имеют важное значение как кормовые растения (хвощ ветвистый - Е. ratnosissimuin, хвощ пятнистый - Е. variegatiun, хвощ зимующий - Е. hiemale). Однако у большинства видов стенки клеток эпидермы инкрустированы кремнеземом, что обесценивает их кормовое значение. Многие виды - трудно искореняемые сорняки пастбищ и полей, особенно с кислыми почвами (хвощ полевой- Е. arvense). Некоторые виды ядовиты для домашних животных (хвощ болотный - Е. palustre, хвощ дубравный - Е. nemorosuin). Хвощи используют как лекарственные растения. Стебли их употребляют вместо наждачной бумаги.
Хвощ полевой (Equisetum arvense) - один из наиболее широко распространенных видов. Это многолетнее травянистое растение. Растет на полях и залежах как сорняк. Его подземная часть представлена корневищем, проникающим в почву на глубину до 1 м. Некоторые укороченные боковые разветвления корневища превращаются в клубни, заполненные запасным крахмалом. В узлах мутовками расположены листовые влагалища и придаточные корни. Корень имеет первичное строение, в коре находятся крупные воздухоносные полости. Надземные побеги двух видов: спороносные, возникающие ранней весной, и стерильные, образующиеся позже и вегетирующие до поздней осени. У других видов хвоща надземные побеги одинаковы. Стерильный побег мутовчато-разветвленный, зеленый, ребристый, в узлах несет сросшиеся в трубку листовые влагалища, заканчивающиеся черными с белой каймой зубцами. Зубцы представляют собой редуцированные листовые пластинки. Органом фотосинтеза служит стебель. На поверхности он имеет однослойную эпидерму с устьичными аппаратами. Под эпидермой расположена кора, состоящая из участков механической и ассимиляционной ткани. Под ними лежит слой основной паренхимы, пронизанный крупными полостями. Заканчивается первичная кора эндодермой. Центральный цилиндр в центре имеет сердцевину из основной паренхимы, клетки которой со временем расходятся, образуя полости. Проводящие ткани собраны в пучки, расположенные по периферии центрального цилиндра. Пучки коллатеральные, закрытые, стебель, как и корень, не способен к вторичному утолщению.
Спороносные побеги более толстые, бурые, без хлорофилла, неветвистые, высотой 15 - 30 см. Они также охвачены в узлах трубчатыми влагалищами с 8 - 9 крупными зубцами. Спороносные колоски образуются на верхушке. После спороношения эти побеги отмирают. Спорангиофор состоит из шестигранного щитка, ножки, прикрепляющей щиток к оси колоска, мешковидных спорангиев, расположенных по нижнему краю щитка. Споры одинаковой величины. Элатеры имеют вид спирально закрученных лент с ложковидными расширениями на концах. На почве из спор вырастают хлорофиллоносные гаметофиты в виде лопастных пластинок, физиологически различные. Одни из них - мужские, с антеридиями, формирующими многожгутиковые сперматозоиды, другие - женские, с архегониями. Оплодотворение связано с водой. Зародыш спорофита не имеет периода покоя.
Таким образом, морфологическая равноспоровость у этого вида сочетается с физиологической разноспоровостыю.
Отдел Папоротниковидные - Polypodiophyta.
Папоротниковидные по древности уступают риниовидным, псилотовидным и плауновидным. Они возникли примерно в одно время с хвощевидными. Но если риниовидные полностью вымерли, а псилотовидные, плауновидные и хвощевидные представлены в современной флоре небольшим числом видов, то папоротниковидные продолжают процветать, хотя и в несколько меньшей степени, чем в прошлые геологические периоды. В настоящее время их насчитывают более 10 тыс. видов. Папоротниковидные распространены по всему земному шару в разнообразных местообитаниях, начиная с тропических лесов и болот и кончая пустынями. Наиболее разнообразно представлены их виды во влажных лесах, где они растут не только на почве, но и на других растениях как эпифиты.
Строение . Спорофиты древних папоротниковидных были древесными растениями с колонновидными неветвящимися стволами, имеющими радиальную симметрию. Затем под влиянием условий умеренного и холодного континентального климата возникли новые виды, приспособленные к геофитному (подземному) образу жизни. Побег у них стал укороченным, травянистым, простертым, дорсивентральным, с придаточными корнями. Современные папоротниковидные - в большинстве случаев многолетние травянистые растения.
Папоротниковидные со всей очевидностью представляют крупнолистную линию эволюции (макрофиллия, или мегафиллия). Листья длительное время нарастают верхушкой. Это позволяет считать, что они образовались путем уплощения теломов, их называют вайями. В большинстве случаев листья совмещают две функции: фотосинтез и спороношение. У некоторых видов верхние вайи специализировались на спороношении, нижние - на фотосинтезе. Большинство видов - равноспоровые, но есть и разноспоровые.
Гаметофит чаще обоеполый. У видов умеренной зоны таллом сердцевидный, у видов тропической зоны - имеет форму нити или ветвистой пластинки. Приспособлен к жизни во влажных условиях. Оплодотворение связано с водой. Гаметофит как бы застыл на уровне водорослевого периода, поэтому, несмотря на то, что спорофит - вполне сухопутное растение, папоротниковидные так и не смогли «завоевать» сушу.
Значение . Папоротниковидные - важный компонент многих растительных сообществ, особенно тропических, субтропических и северных широколиственных лесов. Они - объекты декоративного цветоводства в условиях закрытого и открытого грунта, а также сырье для получения лекарств.
Классификация . Отдел делят на три класса: Первопапоротники, Толстоспорангийные папоротники, Тонкоспорангийные папоротники.
Представители класса Первопапоротники (Primofilipsida) полностью вымерли. Класс Толстоспорангийные папоротники (Eufilipsida) представляет угасающую ветвь эволюции. Общее число видов 150. Класс Тонкоспорангийные папоротники (Leptofilipsida) объединяет подавляющее большинство современных папоротниковидных.
Класс Тонкоспорангийные папоротники - Leptofilipsida.
Общее число видов около 10 тыс. Жизненные формы разнообразны: древовидные, лианы, травянистые эпифиты (влажные тропические леса), многолетние корневищные травы (умеренная и холодная зоны). Подавляющее большинство видов - равноспоровые сухопутные растения. Около 120 видов - разноспоровые, водные и болотные растения.
Щитовник мужской (Dryopteris filix-mas) широко распространен в лиственных лесах, на сырых тенистых местах. Спорофит - многолетнее травянистое растение до 1 м высотой. Побег представлен подземным корневищем. Оно короткое, толстое, черно-бурого цвета, с хорошо выраженным дорсивентральным строением (верхняя поверхность несет черешки листьев, нижняя - тонкие придаточные корни) и заканчивается верхушечной почкой, состоящей из конуса нарастания, окруженного молодыми листьями.
Корневище снаружи покрыто эпидермой. Под ней расположена кора, наружный слой которой состоит из механической ткани. Центр центрального цилиндра занимает сердцевина. Концентрические проводящие пучки расположены по периферии центрального цилиндра. Камбия нет.
Листья крупные. Черешок густо покрыт бурыми пленками. Пластинка в очертании эллиптически-продолговатая, двоякоперисторассеченная. Сегменты первого порядка расположены поочередно, заостренные; сегменты второго порядка с зубчатым краем и тупой верхушкой. Снаружи листа находится эпидерма, клетки которой содержат хлоропласты. На нижней эпидерме много устьичных аппаратов. Мезофилл листа губчатый; проводящие пучки такого же строения, как и в корневище.
На нижней стороне листа вдоль средних жилок сегментов второго порядка расположены группы спорангиев - сории, покрытые сверху пленчатым покрывальцем (индузием) почковидной формы, прикрепленным к выросту листа - плаценте. Спорангий имеет форму чечевицы с длинной ножкой и также прикреплен к плаценте. Оболочка спорангия многоклеточная, однослойная. Среди тонкостенных клеток имеется ряд клеток с подковообразно утолщенными стенками, узкой полосой опоясывающий спорангий. Кольцо этих клеток не смыкается. При подсыхании спорангия клетки кольца сжимаются и происходит разрыв оболочки в тонкой части и освобождение спор. Споры одинаковой величины, имеют овальную форму и бугорчатую поверхность. При образовании спор происходит мейоз.
Из споры, попавшей в благоприятные условия, вырастает гаметофит (заросток). Он имеет вид зеленой округло-сердцевидной пластинки, диаметром до 4 мм, к почве прикреплен ризоидами. На нижней стороне, вблизи от выемки, расположены архегонии с брюшком, погруженным в ткань гаметофита, а в ризоидальной части - округлые антеридии. У равноспоровых папоротников гаметофит обоеполый. Оплодотворение связано с водой. Из зиготы вырастает зародыш спорофита.
Разноспоровые представители класса - преимущественно обитатели тропических и субтропических лесов. На юге европейской части СССР в озерах и заводях рек можно встретить сальвинию плавающую (Salvinia natans). Ее спорофит состоит из разветвленного побега длиной до 15 см, плавающего на поверхности воды. В центре центрального цилиндра стебля находится концентрический проводящий пучок. В коре имеются большие воздушные полости. На стебле мутовками расположены листья. В каждой мутовке три листа, два из них овальные, зеленые, плавающие. Внутри этих листьев также имеются многочисленные воздушные полости. Третий лист каждой мутовки подводный, рассечен на 8 - 12 нитевидных долей, покрытых многоклеточными волосками. Корней нет. Поглощение воды и растворенных в ней минеральных веществ осуществляется всей поверхностью, но главным образом видоизмененными подводными листьями. Сории образуются у основания погруженных листьев. Одни из них содержат микроспорангии с микроспорами, другие - мегаспорангий с мегаспорами, из которых жизнеспособна только одна. Сории осенью погружаются на дно. Весной после сгнивания их покровов спорангии всплывают. Гаметофиты образуются внутри спорангиев. Из микроспор образуются мужские гаметофиты, каждый из которых состоит из двух вегетативных клеток и двух антеридиев. Разрастаясь, мужские гаметофиты разрывают оболочку спорангия и выходят наружу. Из мегаспоры образуется женский гаметофит. Он зеленый, менее редуцирован, чем мужской, верхняя часть его высовывается из мегаспорангия, на ней образуются 3 - 5 архегониев. После оплодотворения из зиготы вырастает небольшой зародыш спорофита, состоящий из стебля и листа.
Таким образом, разноспоровость и тут привела к редукции гаметофитов.
Размножение и распространение которых осуществляется посредством спор, причем споры образуются двумя путями – бесполым и половым. Споровыми растениями являются водоросли , грибы , лишайники и высшие споровые растения (папоротники , плауны , хвощи , мхи , некоторые ископаемые растения).
В процессе эволюции около 400 млн. лет назад от многоклеточных зеленых водорослей возникли риниофиты – первые высшие растения, размножающиеся спорами, которые дали начало всем современным высшим споровым и семенным растениям. Это вымершая группа растений.В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
Высшие споровые растения после выхода на сушу в ходе эволюции претерпевали метаморфозы в двух направлениях. Так сформировались две крупные эволюционные группы – гаплоидная и диплоидная. К первой ветви относятся мхи, у которых лучше развит гаметофит, а спорофит занимает подчиненное положение. К диплоидной ветви принадлежат папоротники, хвощи и плауны. У них гаметофит редуцирован, и выглядит как заросток.
Размножение
Из спор, которые формируют особи бесполого поколения, вырастают особи полового поколения. Они имеют специальные мужские и женские половые органы, в которых развиваются мужские и женские половые клетки (гаметы) – подвижные сперматозоиды и неподвижные яйцеклетки. Для оплодотворения сперматозоид должен попасть во внешнюю среду и оплодотворить яйцеклетку, которая находится внутри женского полового органа. Для перемещения сперматозоида необходима вода. Из оплодотворенной яйцеклетки образуется зародыш. Он прорастает и превращается в особь бесполого поколения, которая размножается спорами.
Грибы, лишайники и водоросли размножаются подвижными зооспорами, спорангиеспорами, неподвижными апланоспорами. Для этих растений характерно половое размножение ооспорами, зигоспорами, вегетативное размножение также посредством спор. Из спор появляется такое же растение, как материнское. Тогда как у высших споровых растений такое растение, как то, что сформировало споры, появляется после сложного процесса оплодотворения.
Спора представляет собой клетку с двойной оболочкой. Споры у определенного вида растений могут быть одинаковыми по размерам или разными. Микроспоры – мелкие, макроспоры – крупные. Микроспоры обычно дают начало мужским заросткам, а макроспоры – женским.
Высшие споровые растения распространены в разных климатических условиях, но большинство из них обитает на влажных участках суши, так как для полового размножения им нужна вода. Однако некоторые виды этих растений встречаются и в пустынях.